消化管平滑筋の生理学と薬理学(Physiology and Pharmacology of Gastrointestinal Smooth Muscles)
- manager
- 久野高義
1. 消化管平滑筋
細長い紡錘形を示す細胞が互いに平行に並んで単位となる小さな束を作る。細胞内にはアクチンとミオシンが存在するが、整然と並んでいることはない。しばしばアクチンと結合してdense body(濃密体)が認められ、これが骨格筋でのZ-band の機能を果たしていると考えられている。また、小胞体の発達も骨格筋と比較して著しく少なく、筋原線維に沿って並ぶことはない。細胞膜表面には平滑筋に特徴的なcaveole(表面小窩)が数多く存在する。
骨格筋との比較
平滑筋 | 骨格筋 | |
|---|---|---|
形態 | 紡錘形で束をなす。 | 円筒形 |
可能な収縮(元の長さの) | 1/4 | 1/2 |
アクチンとミシンの比 | 10-20:1 | 2:1 |
筋小胞体の発達 | 少ない。 | 多くの小胞体が整然と筋原繊維に沿って並ぶ。 |

平滑筋の特徴を表す概略図 |
2. 神経支配と神経・平滑筋伝達
消化管運動は外来神経と内在神経により制御されている。外来神経として、交感神経と副交感神経が支配しているが、これら外来神経の数は内在神経に比べて極めて少ない。腸神経叢にはアミン、アミノ酸、ペプチド系などの中枢神経に存在するほぼ全ての神経伝達物質を含む内在神経が存在する。従って消化管運動は内在神経の活動により主に制御されていると言える。
一方、消化管ではカハール(Cajal)の間質細胞や一部の平滑筋が自発性に活動電位を発生し、消化管運動を生じている。摘出組織でも自発運動を記録することができるのはこのためである。平滑筋は骨格筋に比べ比較的遅い収縮をする。平滑筋収縮を生じる Ca2+ によるシグナル伝達には、細胞外と細胞内ストアーの両方の Ca2+ が用いられ、 Ca2+ 利用のパターンは骨格筋や心筋と異なる。
a、外来神経と内在神経
副交感神経系は迷走神経が支配しているが、下部結腸や直腸では仙部副交感神経も支配している。しかし、例えばネコでの迷走神経遠心性線維の数は少なく、消化管神経叢細胞の約一万分の一と言われる。事実、平滑筋細胞に直接終わる神経線維は少なく消化管平滑筋全体を支配することはない。内在コリン神経を刺激すると平滑筋細胞で興奮性接合部電位(excitatory junction potentials, e.j.ps)がとれる。一方、交感神経系として、下部腸間膜神経節から節後線維がのびているが、殆どが内在神経叢にシナプスを作って終わる。従って、交感神経を刺激しても平滑筋細胞で抑制性接合部電位(inhibitory junction potentials, i.j.ps) がとれることはない。神経叢内において cholinergic neuron の神経終末に終わり、シナプス前抑制により、ACh 遊離を抑制して内在神経の伝達を抑制している。
平滑筋細胞に抑制性の電位を発生させる伝達は、非アドレナリン非コリン(Non-adrenergic non-cholinergic, NANC) 性伝達と呼ばれている。骨格筋では多くの種をこえて運動神経の神経伝達物質は acetylcholine であり、この興奮性支配によってのみ骨格筋の緊張が制御されている。これに較べ、平滑筋には抑制性神経支配が存在し、それが動物種により、腸管部位により著しく異なることが知られている。NANC 性伝達物質として 一酸化窒素、vasoactive intestinal peptide (VIP), neurotensin, ATP などが一部の消化管部位で働いていることが報告されているが、依然として抑制性神経が未知である消化管部位が多く、弛緩機序を含めて詳細は知られていない。
b、自発運動
消化管の平滑筋細胞には、自発性に活動電位を発生する細胞、刺激によって活動電位を発生する細胞、刺激を加えても発生しない細胞とがある。一部の平滑筋細胞で発生する自発性の電位は slow wave と呼ばれ、形や頻度は平滑筋によって異なるが、数 mV から数十 mV の振幅で 1-10 数秒持続する緩やかな電位で 1 分間に 1-20 個発生する。脱分極が閾値に達すると活動電位を発生する。一部、モルモット結腸紐や精管などでは心筋のような spike 型の活動電位を発生する組織もある。tetrodotoxin や atropine が無効でありslow wave は筋原性である。粘膜筋板と輪走筋層の間に存在するCajal cell から発生し、nexus を介して平滑筋組織全体に伝わり腸運動を生じる。活動電位はメディウムの Na+ を除去したり、tetrodotoxinで Na+ 流入を阻害しても影響を受けない。しかし、細胞外の Ca2+ に依存性が高く、 Ca2+ 拮抗薬やMn2+ などで Ca2+ 流入を抑えると活動電位は出なくなり、平滑筋のspike は Ca2+ spike であることが分かる。
c、平滑筋の収縮
(1)Ca2+ によるシグナル伝達
弛緩時の平滑筋細胞内の Ca2+ 濃度は60-200 nM 程度で、収縮時は1-10 mM となる。細胞外液の Ca2+ 濃度は 3mM 程度なので著しい濃度勾配を生じているが、静止時の平滑筋細胞膜は Ca2+ に対し殆ど透過性がない。神経伝達物質の働きにより、細胞内 Ca2+ が高まると収縮機構が活性化され、アクチンとミオシンの相互作用により収縮を生じる。
(2)筋収縮に関わる Ca2+ の供給源
i) Ca2+ channel を通っての外からの Ca2+ 流入
ACh が muscarinic 受容体に結合した時などでは、受容体作動性の非選択性陽イオン channel が開いて Ca2+ が外から流入する。また、平滑筋細胞膜が脱 分極することにより、電位依存性の Ca2+ channel が開いて Ca2+ が流入する。
ii) 細胞内 Ca2+ ストアーである sarcoplasmic reticulum (SR) からの Ca2+ 放出
ACh が muscarinic の m1, m3, m5 受容体に結合した時などでは、phospholipase C (PLC) βの活性化を介して、inositoltrisphosphate (IP3) が産生される。また、tyrosine kinase の活性化により PLCg が IP3 を産生する。IP3 が SR に存在する IP3 受容体を活性化して Ca2+ を放出する。IP3受容体は Ca2+ の存在下にさらに活性化される。また、流入した Ca2+ が SR に存在する ryanodine 受容体を活性化し、 Ca2+ を放出する。この機構は心筋でとくに顕著で、流入した Ca2+ よりもはるかに多量の Ca2+ を SR から放出する。
iii) 細胞質からの Ca2+ の排出抑制
平滑筋細胞膜の plasma membrane Ca2+-ATPase の抑制による細胞外への Ca2+ 排出抑制、あるいは SR に存在するSR Ca2+-ATPase (SERCA 2) の抑制による SR への Ca2+ 取り込み抑制によっても細胞内 Ca2+ は増加する。
(3)平滑筋での細胞内 Ca2+ の増加
消化管平滑筋では、受容体作動性 Ca2+ channel および電位依存性 Ca2+ channel の両方が働いている。また、IP3 受容体およびryanodine 受容体を介して Ca2+ を SR から放出する両方の機序が働いている。それらの働きの重要性は組織とアゴニストの種類により異なる。一般的には、平滑筋収縮の早い第一相にはストアーからの Ca2+ が 利用され、遅い第二相には細胞外からのCa2+ 流入が貢献している。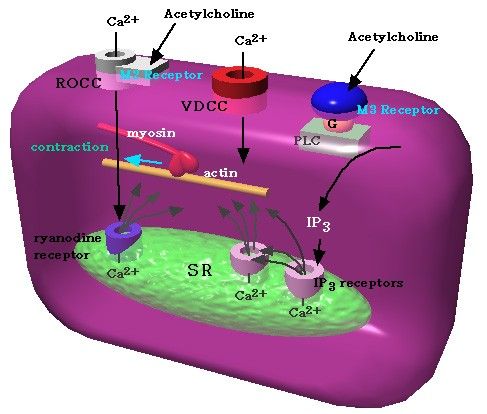
平滑筋細胞でのCa2+動員機構 |
(4)ミオシンのリン酸化と収縮
i) Ca2+ イオンによるミオシン軽鎖キナーゼの活性化
増加した Ca2+ イオンは細胞内カルモジュリンと結合し、ミオシン軽鎖キナーゼを活性化する。その結果、ミオシン軽鎖のリン酸化が起こり、アクチンとミオシン頭部で形成される cross bridge が回転して収縮が生じる。従って、収縮の強さを決めるのは一義的には、ミオシン軽鎖をリン酸化する細胞内 Ca2+ イオン濃度と言える。
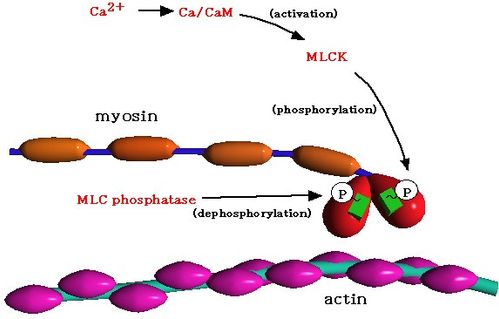
ミシン軽鎖のリン酸化によるcross bridgeの回転 |
ii) ミオシン軽鎖リン酸化量の調節
一定の Ca2+ イオン濃度下で、ミオシン軽鎖のリン酸化を調節する機序が存在する。平滑筋細胞膜に存在する m3 やα1 受容体が活性化されると、低分子量 GTP 結合蛋白質のひとつである RhoA が活性化され、RhoA kinase を介してミオシン軽鎖フォスファターゼがリン酸化され、活性が抑制される。その結果ミオシン軽鎖のリン酸化が増加し収縮反応が出る。これとは別に、PKC が 17kDa 蛋白を介して、やはりミオシン軽鎖フォスファターゼを抑制する系も知られている。これらは、Ca2+ イオン濃度が一定であるのに収縮を生じるので、収縮機構の ‘Ca2+ 感受性増加’とも表現されている。このほか、細胞内 cyclic GMP の増加が PKG を介して、ミオシン軽鎖フォスファターゼを活性化する。また、カルシュウム・カルモヂュリン依存性プロテインキナーゼIIがミオシン軽鎖キナーゼをリン酸化して、ミオシン軽鎖キナーゼを抑制する系も知られている。これらの場合には、Ca2+ イオン濃度が一定であるのに弛緩を生じるので、収縮機構の ‘Ca2+ 感受性低下’とも表現されている。
d、平滑筋の収縮と弛緩のまとめ
平滑筋は体の主な中空臓器に存在しており、消化器、血管、膀胱、子宮などに分布している。その性質は器官ごとに異なっており、収縮と弛緩を引き起こすアゴニストも異なっている。大動脈や気管支の平滑筋は活動電位を発生しない。
収縮系
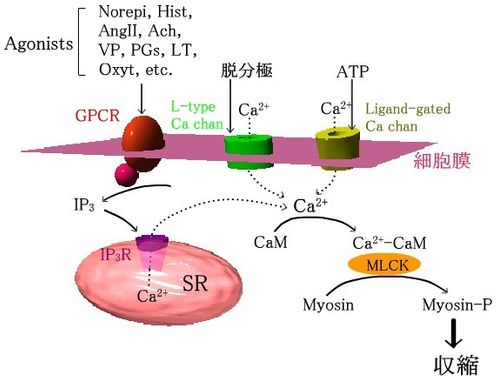
活動電位は、主として直接の機械(伸展)刺激とL型電位依存性Caチャネルにより生じる。これがCaイオンの流入の主な経路である。また、多くの平滑筋には、リガンド依存性イオンチャンネル(P2Xなど)があり、自律神経から遊離されたATPなどに反応し、Caイオンの流入を引き起こす。さらに、さまざまなアゴニストに反応するGPCRが存在し、IP3産生とIP3受容体刺激を介して、SRからCaイオンを遊離する。
|
| 平滑筋の収縮系 GPCR:G蛋白共役型受容体、MLCK:myosin light-chain kinase、CaM:calmodulin、SR:筋小胞体 |
弛緩系
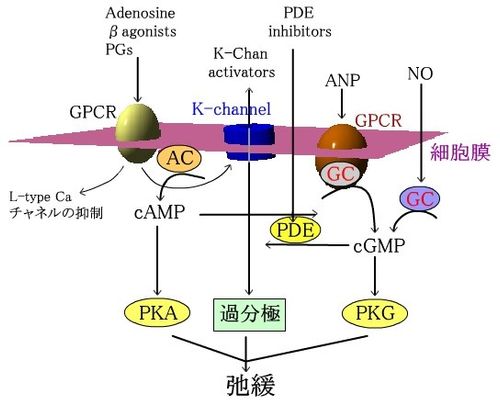
カリウムチャネルと抑制性アゴニストに対するGPCRは、Kイオン流入、cAMP産生、Caチャンネルの調節をしている。ANP受容体は膜結合型GCを、気体のNOは可溶性GCを刺激し、cGMPの産生を引き起こす。PDE阻害薬はcAMPとcGMPの分解を抑制する。cAMPやcGMPは、各protein kinaseを介し、MLCKのリン酸化を引き起こす。リン酸化されたMLCKはCa2+-CaMとの親和性が低下し弛緩を引き起こす。
|
| 平滑筋の弛緩系 NO:nitric oxide、PKA:protein kinase A、PKG:protein kinase G、PDE:phosphodiesterase、ANP:atrial natriuretic peptide、AC:adenylate cyclase、GC:guanylate cyclase 参照:ラング・デール「薬理学」(2012) |
関連サイトの紹介
1、東邦大学メディアセンター 血管平滑筋の収縮と弛緩
(畑、三木)